No correlation between HSPG2 genetic variants and anthropometric
characteristics within an ACL rupture risk modelling study
LethaboRamoshaba
Division of Physiological Sciences, Health through Physical
Activity, Lifestyle and Sport Research Centre (HPALS), Department of Human
Biology, University of Cape Town, Newlands, South Africa
Mary-Jessica NLaguette
Division of Physiological Sciences, Health through Physical
Activity, Lifestyle and Sport Research Centre (HPALS), Department of Human
Biology, University of Cape Town, Newlands, South Africa
International Federation of Sports Medicine (FIMS)
Collaborative Centre of Sports Medicine, University of Cape Town, Newlands,
South Africa
Malcolm Collins
Division of Physiological Sciences, Health through Physical
Activity, Lifestyle and Sport Research Centre (HPALS), Department of Human
Biology, University of Cape Town, Newlands, South Africa
International Federation of Sports Medicine (FIMS)
Collaborative Centre of Sports Medicine, University of Cape Town, Newlands,
South Africa
Alison VSeptember
Division of Physiological Sciences, Health through Physical
Activity, Lifestyle and Sport Research Centre (HPALS), Department of Human
Biology, University of Cape Town, Newlands, South Africa
International Federation of Sports Medicine (FIMS)
Collaborative Centre of Sports Medicine, University of Cape Town, Newlands,
South Africa
BackgroundAnterior cruciate ligament
(ACL) ruptures are common musculoskeletal injuries, influenced by extrinsic
and intrinsic factors such as genetic variations and anthropometric traits.
While these factors contribute to ACL rupture susceptibility, their
interactions are underexplored.
ObjectivesTo investigate the
relationship between HSPG2 variants and anthropometric traits in
participants from an ACL study from Poland and Sweden.
HypothesisGenetic variability within
HSPG2 loci along with height variability may collectively
contribute to ACL rupture susceptibility.
Sample and methodsA genetic
case-control association study was conducted with two cohorts from Poland
and Sweden and a combined cohort. Participants were self-reported Caucasian
and physically active. The combined cohort consisted of 265 asymptomatic
controls (POL-CON=150; SWD-CON=116); 237 ACL rupture cases (POL-ACLR=141;
SWD-ACLR=95) and a subgroup of 135 non-contact ACL ruptures (POL-NON=54;
SWD-NON=79). Participants were genotyped for rs2291826 A>G and rs2291827
G>A and data were analysed using R, with p<0.05.
ResultsStrong correlations were found
between mass and BMI across all cohorts (r=0.78–0.81), suggesting these
traits may influence injury risk. Sex-mass and sex-height correlations were
consistent, with a strong negative correlation between sex and height in the
Swedish cohort (r=-0.75). No positive correlations were found between the
HSPG2 variants and anthropometric traits, except a moderate
negative correlation between rs2291826A>G and height in the Swedish
cohort (r=-0.019, p<0.009), suggesting possible genotype effect on
height.
ConclusionMass and BMI were
highlighted as potential risk factors for ACL rupture. Height-mass
relationships varied by sex and population, suggesting both genetics and
environment impact injury patterns. Further testing of the variants may
clarify their role in ACL injury variability.
Keywords: Anterior cruciate ligament,
body mass index,
HSPG2 ,
Heparan sulphate proteoglycan-2 gene, SNHA,
St Nicholas House analysis
Conflict of interest statement:
There are no conflicts of interest.
Citation: Ramoshaba, L. et al. (2025). No correlation between
HSPG2 genetic variants and anthropometric characteristics within an ACL
rupture risk modelling study. Human Biology and Public Health 1. https://doi.org/10.52905/hbph2025.1.93.
It is important to test the potential relationship between genetic variants and
anthropometric measurements towards identifying potential confounders in a
genetic association study. This study has shown that height is not a confounder
for these variants.
Contents
Introduction
Rupture of
the anterior cruciate ligament (ACL) is one of the most severe and common
musculoskeletal soft tissue knee injuries (Alsayed et al.
2023; Kiapour and Murray 2014). It is
associated with immediate pain, rapid swelling, weakness of the knee, and instability,
which significantly impair mobility (Logerstedt et al.
2018). The incidence rate of ACL ruptures is estimated at 75–80 per 100,000 per
year, especially in physically active adolescents and athletes (15–40 years) (Magnusson et al. 2020). ACL ruptures occur most
frequently in athletes who participate in sporting activities that require sudden
changes in direction, jumps, and twists (Joseph et al.
2013). The lifetime risk of developing knee osteoarthritis as a result of an ACL
injury is 34%, 10–15 years post-injury and surgical repairs (Davis et al. 2021; Suter et al.
2017). It is particularly concerning that these injuries are increasingly
affecting young individuals; consequently, there is a growing number of osteoarthritis
cases appearing at a younger age, which raises further concern (Murray et al. 2000; Lohmander et al.
2007).
Ligaments have a low healing capacity, poor blood supply, and low
cellular content, all of which has been ascribed to contribute to the ACL’s
susceptibility to injury (Kiapour and Murray 2014). As
a result, ACL ruptures not only cause significant direct healthcare costs, but also lead
to indirect costs, such as social and productivity losses and increased absenteeism,
collectively creating a long term economic burden (Griffin
et al. 2006). The burden of ACL injuries is well recognized, however, the
aetiology of ACL rupture remains unclear (Kobayashi et al.
2010). It is known that ACL injuries are multifactorial, meaning that an
interaction between various intrinsic and extrinsic risk factors contribute to the
susceptibility to these injuries (Hewett et al. 2016;
Bittencourt et al. 2016; Meeuwisse 1994). Although research is increasingly focusing on the
contributions of biomechanical (modifiable) risk factors and environmental risk factors
to ACL injuries (Murphy et al. 2003; Griffin et al. 2006), there is growing evidence that an
individual’s genetic predisposition and anthropometric traits contribute to
susceptibility to musculoskeletal soft tissue injuries such as ACL ruptures (Joseph et al. 2013; Magnusson et al. 2020; Rahim et al. 2017; Snaebjörnsson et al. 2019).
More than 80 loci
encoding genes related to components of the extracellular matrix (ECM) have been
identified as being associated with predisposition to ACL rupture (Ribbans et al. 2022). These genes encode proteins which form
structural, non-structural and regulatory components of the ECM (Posthumus et al. 2010; Ribbans et al.
2022; Willard et al. 2018; Feldmann 2022; Feldmann et al.
2022; Dlamini et al. 2023). Recently, Dlamini
et al. (2023) highlighted genetic variants within two
candidate genes, heparan sulphate proteoglycan-2 (HSPG2) and integrin beta 2
(ITGB2), to be associated with ACL rupture susceptibility. They showed that
the G-A-C allele combination between HSPG2 (rs2291826 A>G-rs2291827 G>A)
and ITGB2 (rs2230528 C>T) variants was associated with reduced risk of ACL
rupture as a proxy for gene–gene interaction between these two genes.
The
ITGB2 gene encodes integrin β2 subunit, a heterodimeric transmembrane
receptor primarily involved in leukocyte adhesion and immune responses (Bednarczyk et al. 2020; Huang et al.
2019). The potential contribution of this gene to ACL rupture susceptibility may
arise through the inflammatory pathways and gene–gene interactions (Dlamini et al. 2023). The HSPG2 gene is a multidomain gene
that encodes perlecan, a large basement membrane protein (Gubbiotti et al. 2017). Perlecan belongs to the proteoglycan family, which form
a major component of extracellular matrix (ECM) ground substance (Gubbiotti et al. 2017; Arikawa-Hirasawa 2022). Perlecan plays a multifaceted role in maintaining the
structural and functional integrity of the ECM (Arikawa-Hirasawa 2022). It interacts with various molecules, including
collagens and growth factors such as vascular endothelial growth factor (VEGF) and
fibroblast growth factor (FGF) to maintain ECM stability and support cellular process
(Farach-Carson and Carson 2007; Arikawa-Hirasawa 2022). The expression of HSPG2 contributes
to various developmental and cellular processes including bone and cartilage
development, angiogenesis, inflammatory responses and wound healing (Farach-Carson and Carson 2007; Martinez et al. 2018). Given the role of HSPG2 in skeletal and cartilage
development, it could also contribute to the regulation of height, a known risk factor
for ACL ruptures. While the ITGB2 was identified in previous research, the study
focused on the HSPG2 due to its stronger biological relevance to ligament
integrity and growth regulation.
The exact genetic variants and their interaction
with extrinsic or intrinsic factors have not been interrogated and require exploration.
Some of the intrinsic risk factors include the anthropometric traits such as mass,
height and body mass index (BMI), which have been identified as potential factors
contributing to susceptibility to musculoskeletal soft tissue injuries such as ACL
rupture (Snaebjörnsson et al. 2019; Alsayed et al. 2023). A study by Alsayed et al. (2023) reported that individuals with a BMI greater than 25
kg/m2 were at a greater risk of sustaining a sport-related ACL injury
compared to individuals with a lower BMI. A high BMI may result in increased mechanical
stress on the knee, resulting in knee injury (Alsayed
et al. 2023; Snaebjörnsson et al. 2019). It
has been noted that females have about 9 times greater risk of ACL injury than men,
potentially due to biomechanical, anatomical and hormonal differences (Seneviratne et al. 2004; Bruder et al.
2023). The variability in these factors also has a genetic component, making it
important to identify whether these intrinsic risk factors are also confounders when
trying to characterise the genetic risk susceptibility for a complex phenotype such as
ACL ruptures.
The aim of this study was therefore to investigate the relationship
between genetic variants of HSPG2 and anthropometric factors. It was hypothesised
that genetic variability at the HSPG2 loci, together with variability in height,
may collectively explain the contribution to susceptibility to ACL rupture. In
attempting to characterise the risk factors for ACL rupture susceptibility, it is
important to identify the confounding factors and determine which factors are associated
with each other.
Sample and
Methods
Samples
Participant recruitment
A
case-control genetic association study was conducted comprising two previously recruited
cohorts from Poland and Sweden, respectively, as well as a combined cohort of members
from both populations. Each participant was of self-reported European ancestry and
provided written informed consent to participate in accordance with principles in the
Declaration of Helsinki. All participants in each cohort completed detailed
questionnaires covering demographics, lifestyle habits, occupation details, and sporting
background including type of sporting activity, duration of participation, and
frequency. The questionaries requested medical details related to the ACL injuries, such
as the mechanism of injury, and history of other ligament or tendon injuries. Ethical
approval for this study was obtained from the Human Research Ethics Committee of the
Faculty of Health Science, University of Cape Town (HREC: 026/2023). Approval was also
obtained from each of the respective local ethics committees for the individual cohorts
recruited from the Regional Ethical Review Board in Umeå, Sweden (dnr. 2011‐200‐31 M),
Bioethics Committee for Clinical Research, Regional Medical Chamber, Gdansk, Poland
(KB‐8/16).
The ACL rupture group included individuals older than 18 years at the
time of recruitment. All the participants had a clinical diagnosis of ACL rupture by
physical examination confirmed by magnetic resonance imaging (MRI) or arthroscopy. This
group of participants included injuries sustained through contact and non-contact
mechanisms. Non-contact ACL rupture was analysed as a subgroup of the cases (NON). The
control (CON) group included individuals with no history of an ACL injury or other
ligament or tendon injuries. They regularly participated in similar sports activities as
the affected individuals and were in the same age group. All participants engaged in
regular physical activity, primarily on a recreational level.
Individuals taking
chronic medications or diagnosed with a connective tissue disease or other systemic
disease known to affect connective tissues were excluded. In addition, individuals with
current or previous use of fluoroquinolone antibiotics (within the last 12 months from
the time of recruitment) or previous use of local corticosteroids injections into the
ACL or surrounding areas were excluded, as these factors can have a negative effect on
collagen synthesis and result in matrix degradation thus increasing susceptibility to
ACL injuries.
Participant characteristics
The combined cohort, including participants from the
Polish and Swedish cohorts, consisted of 502 participants. The cohort consisted of 265
asymptomatic control participants (COMB-CON), 237 participants with ACL rupture
(COMB-ACLR), and 135 participants (COMB-NON) who reported sustaining an ACL rupture by a
non-contact mechanism (Table 1). The Polish cohort
consisted of 291 physically active, unrelated participants recruited between 2008 and
2018 from the Galen Orthopaedics Clinic in Poland. This included 150 asymptomatic
control participants (POL-CON), 141 ACL participants (POL-ACLR), and 54 participants who
reported a non-contact mechanism (POL-NON). The male controls (n=112) and ACL rupture
cases (n=102) were recruited from the Polish soccer league. The female cases (n=39) were
recruited from soccer teams and skiing sports. The female controls (n=37) were recruited
from sports clubs and wellness centres (Cięszczyk et al.
2017; Feldmann et al. 2022; Lulińska-Kuklik et al. 2019).
The Swedish cohort consisted of
211 physically active and unrelated participants recruited between 2011 and 2013 from
the orthopedic clinics of two major hospitals in the city of Umeå (in the Vaesterbotten
region) and Luleå (in the Norrbotten region) respectively. This cohort comprised 116
asymptomatic control participants (SWD-CON) with no history of ACL injury, 95
participants (SWD-ACLR) with ACL injury, and 79 participants who reported an ACL rupture
by a non-contact mechanism (SWD-NON). Most of the recruited participants had a
previously described long-term follow-up of the ACL injury (Suijkerbuijk et al. 2019).
Participant sports data were
previously published by Suijkerbuijk
et al. (2019) and Cięszczyk et al. (2017).The type of sport participation was categorised into
non-contact jumping sports, non-contact non-jumping sports, and skiing sports. The
participants in the Polish cohort were matched in terms of sport type, level, and
frequency of participation. Details on the years of sports participation in the Polish
were not available. Most of the participants in the Swedish cohort reported
participating in non-contact jumping sports at a recreational level.
Method
DNA extraction
For the participants in the Polish cohort,
genomic DNA was extracted from oral epithelial cells using a GEN Elute Mammalian Genomic
DNA Miniprep Kit (Sigma; Darmstadt, Germany) according to the manufacturer’s
recommendations. For the Swedish cohort, genomic DNA was extracted from venous blood
using rapid non-ethanol precipitation as previously described by Lahiri and Nurnberger
(1991), with slight modifications from Mokone
et al. (2006).
Candidate gene and genetic
variants selection
The candidate gene HSPG2 was selected because of its
multifunctional role in different biological pathways involved in remodelling the
extracellular matrix (ECM) components and thereby maintaining ECM homeostasis.
Participants were genotyped for the following genetic variants: HSPG2 (rs2291826
A>G, rs2281827 G>A). The genetic variants were selected from a previous Whole
Exome Sequencing (WES) study, which showed that the variants had a difference in allele
frequency distribution of atleast 30% between 10 tendinopathy cases and 10 controls (Gibbon et al. 2018). These genetic variants were also
selected because they reported a minor allele frequency of > 5% in the Caucasian
population, as reported in the ALFA Allele Frequency dataset and 1000 Genomes Project
hosted by the National Centre for Biotechnology Information (NCBI database (http://www.ncbi.nlm.nih.gov/). The functional significance of the
genetic variants remains unknown.
Genotyping
For the standard genotyping protocols, a predesigned TaqMan
polymerase chain reaction (PCR) assay was used to genotype the HSPG2 rs2291826
A>G (Assay ID: C_15966515_10) and HSPG2 rs2291827 G>A (Assay ID:
C__15966517_10) (ThermoFisher Scientific; Waltham, Massachusetts, USA). Real-time
polymerase chain reactions (PCR) were performed for all genetic variants using the Quant
Studio™ 3 Real-Time PCR System (Applied Biosystems; Waltham, Massachusetts, USA). Three
negative controls (no DNA) and five replicate samples (known genotypes) were included on
each 96-walled PCR plate (FG-TCII reaction plate) as a quality control for reliable
genotyping and detection of contamination. Genotypes were confirmed by two independent
investigators. Genotypes were called using the Thermofisher CloudTM Suite
(Thermo Fisher Scientific; Waltham, Massachusetts, USA) with an average call rate of
82.7%. All laboratory work was conducted at the University of Cape Town, at the Health
through Physical Activity, Lifestyle and Sport (HPALS) Research Centre, which is located
at the Sports Science Institute of South Africa (Newlands).
Statistical analysis
The sample size for
the study was calculated using QUANTO version 1.2.4 (http://keck.usc.edu/biostatistics/software/). For the Polish and
Swedish cohorts, a sample size of 118 or more cases was required to detect an OR of 1.8
or higher. For the Swedish cohort, a sample size of 85 cases was required to detect an
odds ratio (OR) of 2.0 or higher. All calculations assumed minor allele frequencies
between 0.2 and 0.5 with a power of 80% and a significance of 5%. The data were analysed
with the R Project for Statistical Computing Version 4.3.1 . The distribution of
continuous data was analysed using the Shapiro-Wilk normality test, and data was
presented as means±standard deviations (Shapiro and Wilk
1965). Non-parametric data was presented as median and interquartile ranges.
Categorical data were compared using the chi-square test or Fisher’s exact test (sample
size <10) and presented as percentages. One-way analysis of variance (ANOVA) was used
for continuous parametric data, while the Mann-Whitney U or Kruskal-Wallis test was used
for non-parametric data.
The R packages genetics, version 1.3.8.1.3 (González et al. 2007) and SNPassoc version 2.1.0 (Schaid et al. 2012) were used to determine the
differences in genotypes and allele frequencies between the groups and to calculate the
probabilities of Hardy-Weinberg equilibrium (HWE) for the variants and linkage
disequilibrium between the HSPG2 genetic variants. The HSPG2: rs2291826
A>G, rs2291827G>A variants were in a moderate positive LD (D'=0.703), with a
significant non-random association (X2=912.35; p<0.001) of the alleles at the two
loci. GraphPad Prism version 10.1.2 (GraphPad Software., Boston, Massachusetts, USA) was
used to visualize genotype and allele frequencies between the different populations. The
St Nicholas House Analysis (SNHA) was used to visually represent the relationship
between different variables such as genetic variables, demographic and anthropometric
factors and phenotypic traits (Hermanussen et al.
2021).
Results
Participant characteristics
In
the combined cohort, the COMB groups (CON vs. ACLR), were covaried for sex and country
of recruitment Table 1. The COMB-ACLR group (33.8 ±
11.7 years, n=235) was significantly older than the COMB-CON group (p= 0.036, 31.3 ±
14.2 years, n = 263). The COMB-ACLR group also had a significantly higher mass (p =
0.016, 75.4 ± 14.2 kg, n = 219) compared to the COMB-CON group (72.4 ± 12.7 kg, n=256);
however, after the height variable was covaried for sex, the significance was no longer
noted. No significant differences were noted for the COMB-NON subgroup.
For the
individual cohorts, Polish cohort had younger participants compared to the Swedish
cohort participants. The POL-CON group (21.0 ± 1.8 years, n = 149) was significantly
younger than the POL-ACLR group (p < 0.001, 31.6 ± 10.0 years, n = 142) and the
POL-NON subgroup (p < 0.001, 30.7 ± 9.9 years, n = 56). However, for the Swedish
cohort, the SWD-CON group (44.7 ± 11.9 years, n = 114) was significantly older than the
SWD-ACLR group (p < 0.001, 37.1 ± 13.3 years, n = 93) and SWD-NON subgroup (p <
0.001, 36.5 ± 13.7 years, n = 78) (Table 1).
Table 1 Participant’s descriptive
characteristics in the combined cohort (COMB) (male and female) as well as
individual cohorts (Poland, Sweden), for the control (CON), anterior cruciate
ligament ruptures groups (ACLR), and non-contact mechanism anterior cruciate
ligament rupture subgroup (ACL-NON).
COMB-CON
COMB-ACLR
p values
COMB-NON
p values
N
265
237
135
Age(years)
31.3±14.2 (263)
33.8±11.7 (235)
0.036 (0.002)B
34.0±12.6 (134)
0.054
Sex (%male)
57.4 (152)
62.0 (147)
0.331
66.7 (90)
0.096
Height (cm)
175.6±10.2 (257)
175.6±9.5 (224)
0.985
176.3±9.3 (127)
0.503
Mass (kg)
72.4±12.7 (256)
75.4±14.2 (219)
0.016 (0.222)C
75.1±15.0 (122)
0.072
BMI
(kg.m-2)
22.7±4.9 (249)
23.41±6.1 (219)
0.186
22.9±6.5 (126)
0.738
POL-CON
POL-ACLR
p
values
POL-NON
p
values
N
149
141
54
Age (years)
21.0±1.8 (149)
31.6±10.0 (142)
<0.001
30.7±9.9 (56)
<0.001
Sex (%male)
75.2 (112)
71.8 (102)
0.519
85.7 (48)
0.113
Height (cm)
178.0±9.7 (149)
177.3±9.6 (139)
0.545
180.0±8.8 (56)
0.176
Mass (kg)
72.6±12.0 (149)
79.0±14.8 (140)
<0.001 (0.086)A
81.4±16.7 (56)
0.000 (0.019)A
BMI
(kg.m-2)
22.8±2.5 (149)
25.0±4.0 (137)
<0.001 (0.078)A
25.1±5.1 (56)
<0.001 (0.096)A
SWD-CON
SWD-ACLR
p
values
SWD
-NON
p
values
N
116
95
79
Age (years)
44.7±11.9 (114)
37.1±13.3 (93)
<0.001
36.5±13.7 (78)
<0.001
Sex (%male)
34.5 (40)
47.4 (45)
0.059
53.2 (42)
0.010 (0.002)A
Height (cm)
172.3±10.1 (108)
172.8±8.6 (85)
0.730
173.4±8.6 (71)
0.435
Mass (kg)
72.1±13.8 (107)
69.1±10.8 (79)
0.113
69.8±10.9 (66)
0.252
BMI
(kg.m-2)
23.7±2.6 (44)
22.9±2.6 (73)
0.125
22.9±2.6 (62)
0.134
Values are expressed as mean±standard deviation, and sex is represented as a
percentage.
p-values in bold typeset indicate significance (p<0.05).
p-values in parentheses are adjusted for the following variables: A Age, B
country of recruitment, and C Sex.
The Polish and Swedish groups were similar in height. However, with regards to
mass, the Polish cohort had a significantly higher mass and BMI. The POL-ACLR group had
a significantly higher mass (p < 0.001, 79.0 ± 14.8 n = 140) and a higher BMI (p <
0.001, 25.0 ± 4.0 kg.m-2, n = 137) than the POL-CON group. Similarly, the high mass and
BMI was noted in the POL-NON subgroup (mass: p < 0.001, 81.4 ± 16.7 kg, n = 56; BMI:
p < 0.001, 25.1 ± 5.1 kg.m-2, n =56). After the mass and BMI were covaried for age,
the significance remained only for the POL-NON subgroup (Table 1).
No significant genotype effect on any of the descriptive
variables was noted for the combined cohort (Table 2).
For the individual cohorts, genotype effect for height was only noted for the Swedish
cohort for the HSPG2 rs2291826 A>G variant distribution where the A/A and A/G
genotype was frequently noted in the distribution (p = 0.009) (Table S4).
Table 2 Genotype effect on descriptive
characteristics for the HSPG2: rs2291826 A>G, and rs2291827 G>A
genetic variants in the combined cohort (COMB).
Age (yrs.)
p value
Height (cm)
p value
Mass (kg)
p value
BMI (kg/m2)
p value
Sex (%M)
p value
HSPG2 rs2291826 A/G
A/A
31.8 ±12.4 (267)
0.199
175.7±9.97 (258)
0.266
74.2 ±14.0 (254)
0.083
23.1±5.8 (250)
0.345
61.0 (164)
0.705
A/G
32.6 ±13.7 (192)
175.1±9.9 (186)
72.6±12.7 (183)
22.8±5.3 (182)
57.7 (112)
G/G
36.1±15.1 (33)
178.2±9.2 (31)
78.2±14.7 (32)
24.3±3.2 (30)
63.6 (21)
HSPG2 rs2291827 G/A
G/G
32.6±12.5 (330)
0.349
175.8±10.5 (320)
0.666
74.2 ±13.9 (313)
0.299
22.9±5.9 (310)
0.458
60.1 (200)
0.291
G/A
31.6 ±13.9 (151)
175.2±8.8 (144)
72.7±12.2 (144)
23.3±4.3 (142)
60.3 (91)
A/A
36.9 ±15.5 (12)
173.9±5.6 (12)
77.9±19.7 (13)
24.8±4.5 (11)
38.5 (5)
All variables except sex are expressed as mean±standard deviation with the number
of participants presented in parentheses.
Sex is expressed as percentages with the number of participants written in
parentheses.
HSPG2: Heparan sulphate proteoglycan 2.
Genotype and
allele frequency
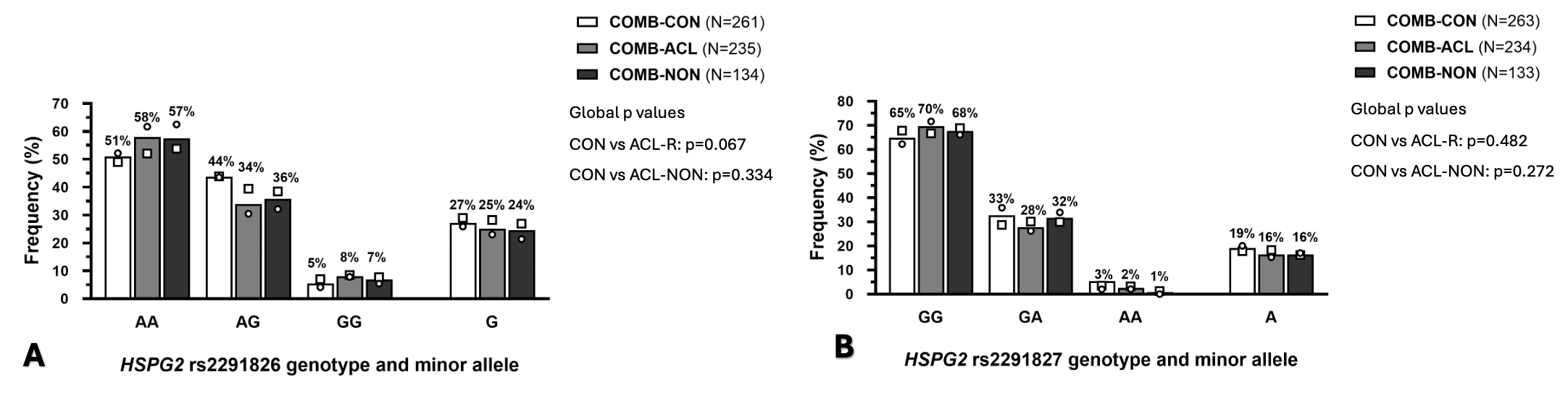
There were no significant differences in the distribution of
genotype and allele frequencies for the HSPG2 rs2291826 A>G variant (COMB-CON
vs. COMB-ACLR: p = 0.067, and COMB-CON vs. COMB-NON: p = 0.334) between the groups
(Figure 1A). Similar findings were noted for the
HSPG2 rs2291827 G/A variant: (COMB-CON vs. COMB-ACLR: p=0.482, and COMB-CON
vs. COMB-NON: p=0.272) (Figure 1B). No significant
differences in either the genotype or allele frequency distributions for the
HSPG2 rs2291826 A>G (Figure 2A) and
rs2291827 G>A (Figure 2B) variant were noted in the
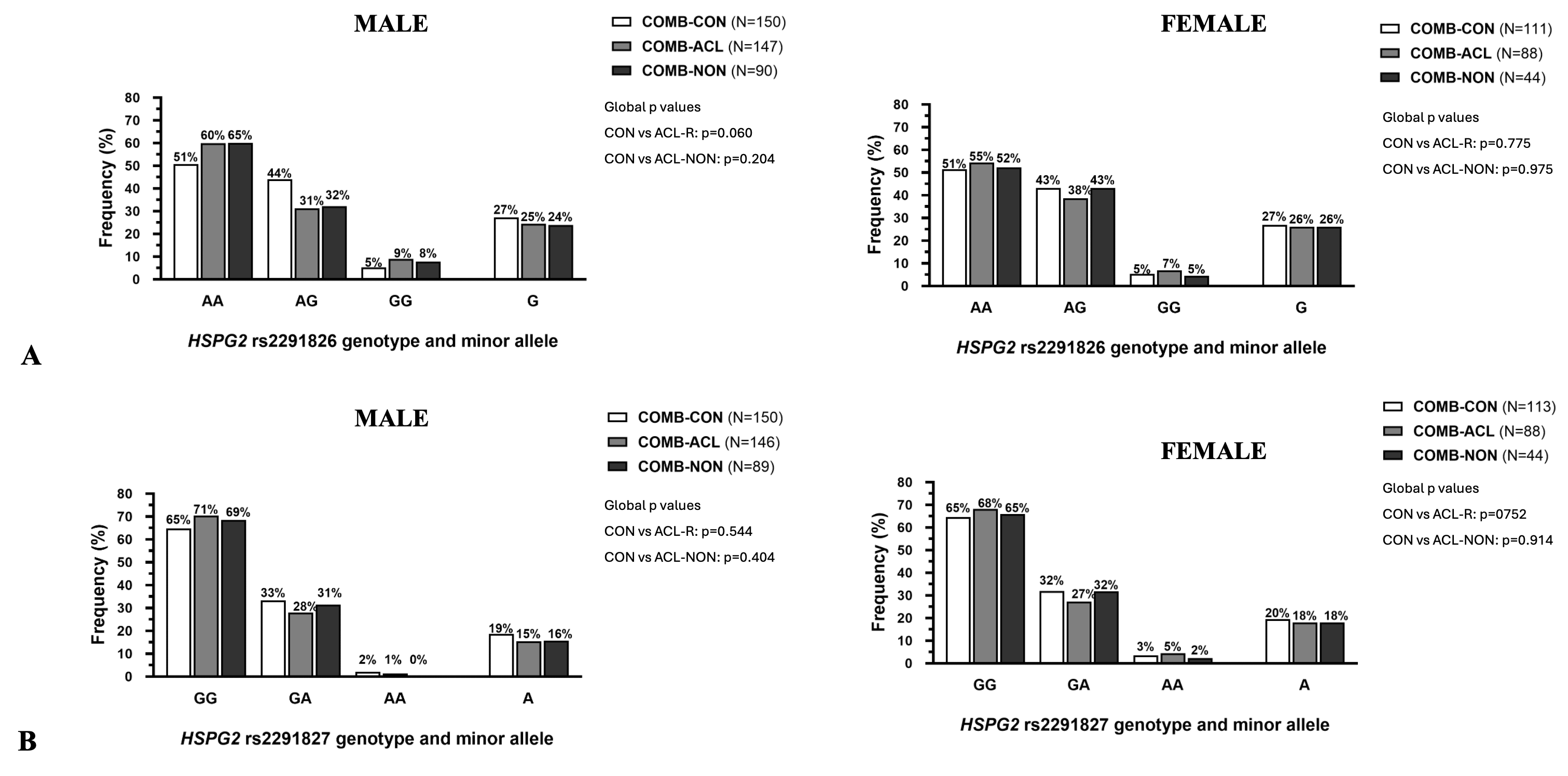
combined cohort when only males or females were examined.
Figure 1 Genotype and allele frequency
distributions of (A) HSPG2 rs2291826 A/G and (B) HSPG2
rs2291827 G/A variant for the combined cohort control group (COMB-CON: white
bars), combined anterior cruciate ligament rupture group (COMB-ACLR: light
grey bars), and ACL rupture by non-contact mechanism subgroup (COMB-NON:
dark grey bars). The differences in genotype and allele distributions within
each individual cohort are annotated as follows: Poland (circle) and Sweden
(square blocks). Details of the HWE and AIC values can be found in Table S5.
Figure 2 Genotype and allele frequency
distributions of (A) HSPG2 rs2291826 A/G and (B) HSPG2
rs2291827 G/A variant for the combined cohort control group (COMB-CON: white
bars), combined anterior cruciate ligament rupture group (COMB-ACLR: light
grey bars), and ACL rupture by non-contact mechanism subgroup (COMB-NON:
dark grey bars). Details of the HWE and AIC values can be found in Table S4 (males) and Table S5 (females).
Relationship
between Genetic variants and Anthropometric Factors
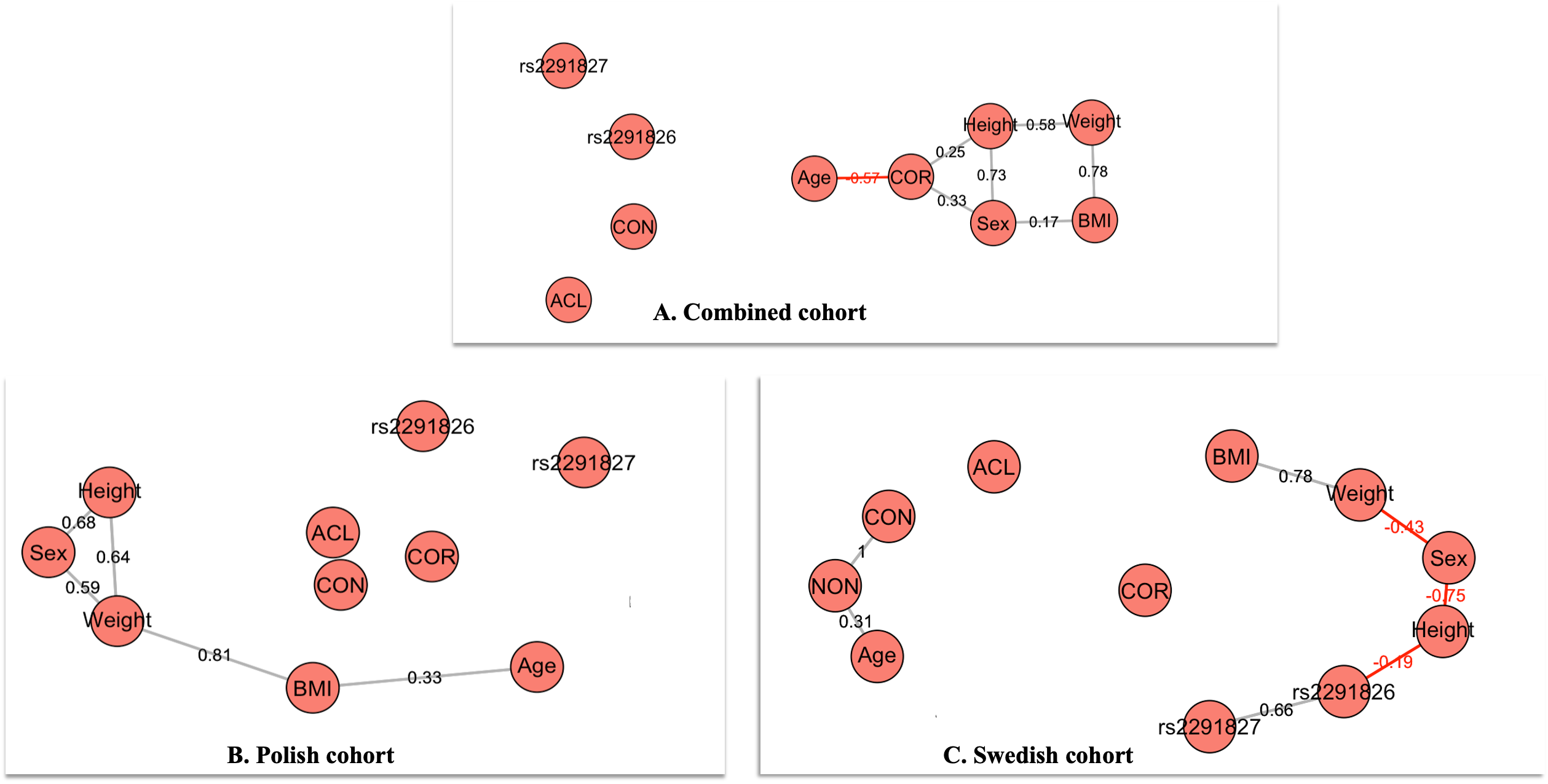
A positive correlation in
mass and BMI was found in all individual cohorts; COMB (r = 0.78), (POL: r = 0.81), and
SWD: r = 0.78) respectively Figure
3. BMI was consistently influenced by mass in all the cohorts and height was
similarly distributed between cases and controls between the cohorts. There appears to
be a consistent correlation between sex with mass and height in all cohorts, with a
negative correlation in the Swedish cohort (r = -0.43) (Figure 3C).
The combined (r = 0.73) and Polish
cohorts (r = 0.68) had a strong positive correlation between sex and height cohort while
a strong negative correlation was noted in the Swedish cohort (r = -0.75), which may be
ascribed to the sex-specific differences in height noted between the cohorts. A positive
correlation was found between COR and height (r = 0.25) in the COMB cohort, which could
possibly be influenced by the height of the Polish participants. No positive correlation
between the HSPG2: rs2291826 A>G and rs2291827 G>A was noted with the
combined and Polish cohorts. However, a negative correlation of the HSPG2
rs2261826 A/G with height (r = -0.19) was found in the Swedish cohort. A positive
correlation was found between the two HSPG2 rs2291826 A>G and rs2291827 G>A
(r = 0.66) variants, which is expected, considering that the genetic variants are in a
moderate positive linkage disequilibrium (D' = 0.703).
Figure 3 St Nicholas House analysis (SNHA)
plot for the combined cohort and individual populations. The grey lines
indicate a positive correlation and the red line a negative correlation
(COR: country of recruitment, BMI: body mass index).
Discussion
The study aimed to
investigate the relationship between genetic variants of HSPG2: rs2291826 A>G
and rs2291827 G>A and anthropometric traits with the risk of ACL rupture in a
combined group of participants recruited from Poland and Sweden. It was hypothesised
that genetic variability at the HSPG2 loci together with variability in height may
collectively explain the contribution to susceptibility to ACL rupture. There was no
significant association between anthropometric traits and the risk of ACL rupture. A
genotype effect for height was found in the Swedish cohort for the HSPG2
rs2291826 A>G variant, where genotypes A/A and A/G were frequently noted in the
distribution. The SNHA plot also showed a negative correlation between genetic variant
HSPG2 rs2291826 A>G and height in the Swedish cohort, suggesting that
individuals with genotypes A/A and A/G tend to be shorter.
In the combined cohort,
significant associations in age and mass were found between the ACL rupture group and
the control groups. The COMB-ACLR group was significantly older (p=0.036) and had a
higher mass (p = 0.016) compared to the CON groups. However, when mass was adjusted for
sex, significance was removed, suggesting that sex and mass may be confounding factors.
The results are consistent with previous studies that emphasize the influence of age and
mass on the risk of ACL rupture (Hurd et al. 2008; Adouni et al. 2024). Excessive mass can increase
mechanical stress on the knee joint and change the kinematics of the joint, which can
lead to knee injuries such as ACL ruptures (Adouni et al.
2024). In the individual cohorts, the Polish cohort was significantly younger
compared to the Swedish cohort, with the POL-ACLR group having a higher mass and BMI
compared to the CON group (p<0.001) (including non-contact ACL rupture). After
adjusting for age, the significance of higher BMI was only maintained for the POL-NON
subgroup. This suggests that age may play a role in the mass and BMI differences
observed in the Polish cohort. Similar trends have been observed in previous studies
where younger people tended to participate in high risk sporting activities, possibly
predisposing them to injury (Motififard et al. 2024;
Snaebjörnsson et al. 2019).
The SNHA
revealed a strong positive correlation between mass and BMI in all cohorts: COMB cohort
(r = 0.78), the SWD cohort (r = 0.78), and an even stronger correlation in the Polish
cohort (r = 0.81) (Figure 3). This is to be expected as
BMI is a mass-related index and the heights between the cases and controls were similar.
However, the broader associations involving anthropometric traits such as mass and BMI,
suggests these factors may influence ACL injury risk (Kızılgöz et al. 2019; Snaebjörnsson et al.
2019; Alsayed et al. 2023). A higher BMI can
lead to excessive stress on the knee joint and can place additional strain on ligaments,
cartilage, and tendons as well as decreased neuromuscular control around the knee
joints, potentially increasing the risk of ACL ruptures (Widmyer et al. 2013; Snaebjörnsson et al.
2019; Motififard et al. 2024). When exploring
the relationship between sex, height and mass, a negative correlation was found in the
Swedish cohort, especially between sex and height (r = -0.75) and sex and mass (r =
-0.43). However, a strong positive correlation between sex and height was also found in
the combined cohort and the Polish cohort. The differences in the patterns could reflect
the sex-specific differences in anthropometric traits influenced by the differences in
the average height of individuals recruited in the two countries. In particular, males
in the Swedish cohort were shorter and weighed less than males in the Polish cohort,
which may have influenced the direction and strength of the observed
correlation.
Although a strong association was found between anthropometric traits
and the risk of ACL rupture, no correlation was found between the genetic variants
HSPG2: rs2291826 A>G, rs2291827 G>A and anthropometric traits in the
combined as well as in the Polish cohort. However, a moderate negative correlation
between genetic variant HSPG2 rs2291826 A/G and height was found in the Swedish
cohort (r=-0.019) (Figure 3C). A
genotype effect on height related to HSPG2 rs2291826 A>G was also noted in the
Swedish cohort. Although the effect size is small, the results suggest that individuals
with an A/A and A/G genotype of HSPG2 rs2291826 A>G may be shorter. The
findings are important because short individuals who weigh more (or have a higher BMI)
may be at increased risk of ACL rupture due to the biomechanical load on the knee joint
relative to stature. Although the HSPG2 rs2291826 variant may not directly influence ACL
rupture risk, but it could have an indirect modifying effect on height. The HSPG2
gene encodes perlecan, a multifunctional protein found in the basement membrane (Iozzo 2005; Farach-Carson and Carson 2007). Perlecan is mainly expressed in the cartilage
matrix and plays an important role in regulating the availability and activities of
growth factors such as fibroblast growth factors (FGF) and bone morphogenetic proteins
(BMPs) (Iozzo 2005). These growth factors play an
important role in the proliferation and differentiation of chondrocytes, an important
process for skeletal development and endochondral ossification (Hayes et al. 2022). The exact biological mechanisms are still
unclear and should be investigated in a larger cohort.
The study had several
limitations, including the sample size of each cohort, in which controls and cases were
not matched by sex and mass. In addition, the cohorts were not matched by age. The
Polish cohort had younger participants, while the Swedish cohort had older participants.
In addition, positive correlations of COR with height and sex could indicate possible
recruitment bias or population-specific characteristics in the cohort, such as regional
differences in height and sex distribution, which could have an influence on the
analysis if not considered. Sport participation data were self-reported, and
participants were not matched for participation in contact and non-contact
sports.
Conclusion
The combined
analysis of participant characteristics and the St. Nicholas house plot highlighted the
role of anthropometric traits, particularly mass and BMI. The positive correlations
between mass and BMI across all cohorts emphasize the importance of mass as a key factor
in the risk of ACL rupture, while the sex-and population-specific differences in the
relationships between height and mass suggest that ACL injury patterns are influenced by
both genetic and environmental factors. There was no positive association between the
genetic variants tested and anthropometric characteristics of participants. For this
reason, it is hypothesised that for these two genetic variants tested, the variability
in ACL risk is most likely not largely influenced by a genetic component of these
measurements as influenced by HSPG2. The variability observed in the risk to ACL
rupture can therefore be tested using the two variants HSPG2 rs2291826 A>G and
rs2291827 G
Appendix
Table S1 Genotype effects on descriptive
characteristics for the HSPG2 genetic variants: rs2291826 A>G and
rs2291827 G>A, in the Polish cohort.
Age (yrs.)
p value
Height (cm)
p value
Weight (kg)
p value
BMI
(kg/m2)
p value
Sex (% M)
p value
HSPG2 rs2291826 A/G
A/A
26.6±8.5 (164)
0.196
176.8±10.0 (161)
0.218
75.6±15.1 (162)
0.907
24.0±3.8 (159)
0.652
70.7 (116)
0.352
A/G
25.2±8.9 (107)
178.7±9.1 (107)
75.9±12.1 (107)
23.7±2.8 (107)
78.5 (84)
G/G
28.8±1.9 (17)
179.4±10.1 (17)
77.2±10.4 (17)
24.0±2.8 (7)
76.5 (13)
HSPG2 rs2291827 G/A
G/G
26.6±8.3(193)
0.483
177.6±10.3 (191)
0.953
76.3±14.9 (191)
0.663
24.0±3.7 (189)
0.594
72.0 (139)
0.547
G/A
25.2±8.9 (107)
178.7±9.1 (107)
75.9±12.1(107)
23.7±2.8 (107)
77.8 (70)
A/A
27.3±9.2 (6)
176.5±6.1 (6)
73.5±10.7(6)
23.5±2.8 (6)
66.7 (4)
All variables except sex are expressed as mean±standard deviation with the
number of participants presented in parentheses.
Sex is expressed as percentages with the number of participants written in
parentheses.
HSPG2: Heparan sulphate proteoglycan 2, and POL: Poland.
Table S2 Genotype effects on descriptive
characteristics for the HSPG2 genetic variants: rs2291826 A>G, and
rs2291827 G>A in the Swedish cohort.
Age (yrs.)
p value
Height (cm)
p value
Weight (kg)
p value
BMI
(kg/m2)
p value
Sex (%M)
p value
HSPG2 rs2291826 A/G
A/A
40.1±13.0 (103)
0.448
173.8±9.8 (97)
0.009
71.6±11.3 (92)
0.094
21.4±7.9 (91)
0.303
45.7 (48)
0.117
A/G
41.9±13.0 (85)
170.2±8.9 (79)
67.9±12.1(76)
21.4±7.4 (75)
32 .2(28)
G/G
43.9±14.4(16)
176.8±8.1 (14)
79.3±18.7(15)
24.7±3.8 (13)
50.0 (8)
HSPG2 rs2291827 G/A
G/G
41.0±12.7 (137)
0.587
173.2±10.3 (129)
0.324
71.0±11.4 (122)
0.755
21.1±8.0 (121)
0.826
43.6 (61)
0.177
G/A
42.9±12.9 (95)
172.6±9.3 (90)
70.5±12.9 (90)
22.3±6.34(83)
34.4 (21)
A/A
46.5±15.0 (6)
171.3±3.9 (6)
81.6±25.5 (7)
26.3±5.9 (5)
14 .3(1)
Table S3 Genotype and minor allele frequency
distributions, p-values, Hardy-Weinberg exact test (HWE), and Akaike
information criterion (AIC) for HSPG2: rs2261826 A>G and rs2291827 G>A
genetic variant in the combined cohort (males and females). ).
COMBINED
MALE + FEMALE
CON %(n)
ACL-R %(n)
p value
AIC
NON %(n)
p value
AIC
HSPG2 rs2291826 (A/G)
n
261
235
134
AA
51.0 (133)
57.9 (136)
0.067
686.8
57.5 (77)
0.334
510.7
AG
43.7 (114)
34.0 (80)
35.8 (48)
GG
5.4 (14)
8.1(19)
6.7 (9)
G
27.2 (143)
25.1 (118)
0.498
24.6 (66)
0.488
HWE
0.118
0.164
0.647
HSPG2 rs2291827 (G/A)
n
263
234
133
GG
64.6 (170)
69.7 (163)
0.482
691.8
67.7 (90)
0.272
509.7
GA
32.7 (86)
27.8 (65)
31.6 (42)
AA
2.7 (7)
2.6 (6)
0.8 (1)
A
19.1 (100)
16.5 (77)
0.332
16.5 (44)
0.451
HWE
0.423
1.000
0.125
Genotype and allele frequencies are expressed as a percentage with the number
of participants (n) in parentheses.
COMB-CON vs COMB-ACLR and COMB-CON vs COMB-NON p values are unadjusted.
P-values in bold typeset indicate significance p< 0.05.
Table S4 Genotype and minor allele frequency
distributions, p-values, Hardy-Weinberg exact test (HWE), and Akaike
information criterion (AIC) for HSPG2: rs2261826 A>G and rs2291827 G>A
genetic variant in the combined cohort (only males). ).
COMBINED
MALE
CON %(n)
ACL-R %(n)
p value
AIC
NON %(n)
p value
AIC
HSPG2 rs2291826 (A/G)
n
150
147
90
AA
50.7 (76)
59.9 (88)
0.060
412.1
60.0 (54)
321.3
AG
44.0 (66)
31.3 (46)
32.2 (29)
0.204
GG
5.3 (8)
8.8 (13)
7.8 (7)
G
27.3 (82)
24.5 (72)
0.486
23.9 (43)
0.468
HWE
0.400
0.128
0.218
HSPG2 rs2291827 (G/A)
n
150
146
89
GG
64.7 (97)
70.5 (103)
0.544
415.1
68.5 (61)
0.404 0.404 0.403
318.6
GA
33.3 (50)
28.1 (41)
31.5 (28)
AA
2.0 (3)
1.4 (2)
0.0 (0)
A
18.7 (56)
15.4 (45)
0.345
15.7 (28)
HWE
0.411
0.534
0.579
Genotype and allele frequencies are expressed as a percentage with the number
of participants (n) in parentheses.
COMB-CON vs COMB-ACLR and COMB-CON vs COMB-NON p values are unadjusted.
P-values in bold typeset indicate significance p< 0.05.
Table S5 Genotype and minor allele frequency
distributions, p-values, Hardy-Weinberg exact test (HWE), and Akaike
information criterion (AIC) for HSPG2: rs2261826 A>G and rs2291827 G>A
genetic variant in the combined cohort (only females).
COMBINED
FEMALE
CON %(n)
ACL-R %(n)
p value
AIC
NON %(n)
p value
AIC
HSPG2 rs2291826 (A/G)
n
111
88
44
AA
51.4 (57)
54.5 (48)
0.775
278.7
52.3 (23)
0.975
190.9
AG
43.2 (48)
38.6 (34)
43.2 (19)
GG
5.4 (6)
6.8 (6)
4.5 (2)
G
27.0 (60)
26.1 (46)
0.932
26.1 (23)
0.986
HWE
0.664
0.750
0.382
HSPG2 rs2291827 (G/A)
n
113
88
44
GG
64.6 (73)
68.2 (60)
0.752
281.0
65.9 (29)
0.914
192.1
GA
31.9 (36)
27.3 (24)
31.8 (14)
AA
3.5 (4)
4.5 (4)
2.3 (1)
A
19.5 (44)
18.2 (32)
0.843
18.2 (16)
0.920
HWE
1.000
0.671
0.288
Genotype and allele frequencies are expressed as a percentage with the number
of participants (n) in parentheses.
COMB-CON vs COMB-ACLR and COMB-CON vs COMB-NON p values are unadjusted.
P-values in bold typeset indicate significance p< 0.05.
Acknowledgements
The statistical analysis and manuscript were the result of
participating in the 7th International Summer School (University of Potsdam)
funded by KoUP University of Potsdam, in Gülpe, Brandenburg, Germany from the
3rd of July to 12th July 2024. I extend great thanks to PD Dr.
Christiane Scheffler, Dr. Detlef Groth, Prof. Michael Hermanussen and Prof. Barry Bogin
for accommodation and patience, their expertise and adventures they created to make the
summer schools enjoyable. I extend utmost appreciation to my supervisors, Professor
Alison September, Professor Malcolm Collins and Dr Mary-Jane Laguette, for the
opportunity to participate in the Summer School and for their invaluable support and
guidance during my MSc journey.
Funding statement
The National Research Foundation
(NRF) supported L.Ramoshaba. The research study was supported by the South African
Medical Research Council (SAMRC) through its Division of Research Capacity Development
under the Mid-Career Scientist Programme, which is funded by the South African National
Department of Health (SANDH).
References
Adouni, M./Aydelik, H./Faisal, T. R./Hajji, R.
(2024). The effect of body weight on the knee joint biomechanics based on
subject-specific finite element-musculoskeletal approach. Scientific Reports
14 (1), 13777. https://doi.org/10.1038/s41598-024-63745-x.
Alsayed, H.
N./Alkhateeb, M. A./Aldossary, A. A./Houbani, K. M./Aljamaan, Y.
M./Alrashidi, Y. A. (2023). Risk of anterior cruciate ligament injury in
population with elevated body mass index. Medicinski Glasnik 20 (1). https://doi.org/10.17392/1517-22.
Arikawa-Hirasawa, E. (2022). Impact of the heparan
sulfate proteoglycan perlecan on human disease and health. American Journal
of Physiology. Cell Physiology 322 (6), C1117-C1122. https://doi.org/10.1152/ajpcell.00113.2022.
Bednarczyk, M./Stege,
H./Grabbe, S./Bros, M. (2020). β2 Integrins-Multi-Functional Leukocyte
Receptors in Health and Disease. International Journal of Molecular Sciences
21 (4). https://doi.org/10.3390/ijms21041402.
Bittencourt, N. F. N./Meeuwisse, W. H./Mendonça, L.
D./Nettel-Aguirre, A./Ocarino, J. M./Fonseca, S. T. (2016). Complex systems
approach for sports injuries: moving from risk factor identification to
injury pattern recognition-narrative review and new concept. British Journal
of Sports Medicine 50 (21), 1309–1314. https://doi.org/10.1136/bjsports-2015-095850.
Bruder, A.
M./Culvenor, A. G./King, M. G./Haberfield, M./Roughead, E. A./Mastwyk,
J./Kemp, J. L./Ferraz Pazzinatto, M./West, T. J./Coburn, S. L./Cowan, S.
M./Ezzat, A. M./To, L./Chilman, K./Couch, J. L./Whittaker, J. L./Crossley,
K. M. (2023). Let's talk about sex (and gender) after ACL injury: a
systematic review and meta-analysis of self-reported activity and
knee-related outcomes. British Journal of Sports Medicine 57 (10), 602–610.
https://doi.org/10.1136/bjsports-2022-106099.
Cięszczyk,
P./Willard, K./Gronek, P./Zmijewski, P./Trybek, G./Gronek, J./Weber-Rajek,
M./Stastny, P./Petr, M./Lulińska-Kuklik, E./Ficek, K./Kemeryte-Riaubiene,
E./Maculewicz, E./September, A. V. (2017). Are genes encoding proteoglycans
really associated with the risk of anterior cruciate ligament rupture?
Biology of Sport 34 (2), 97–103. https://doi.org/10.5114/biolsport.2017.64582.
Davis, A. M./Wong,
R./Steinhart, K./Cruz, L./Cudmore, D./Dwyer, T./Li, L./Marks, P./McGlasson,
R./Urquhart, N./Wilson, J. A./Nimmon, L./Ogilvie-Harris, D./Chahal, J.
(2021). Development of an intervention to manage knee osteoarthritis risk
and symptoms following anterior cruciate ligament injury. Osteoarthritis and
Cartilage 29 (12), 1654–1665. https://doi.org/10.1016/j.joca.2021.08.011.
Dlamini, S.
B./Saunders, C. J./Laguette, M.-J. N./Gibbon, A./Gamieldien, J./Collins,
M./September, A. V. (2023). Application of an in silico approach identifies
a genetic locus within ITGB2, and its interactions with HSPG2 and FGF9, to
be associated with anterior cruciate ligament rupture risk. European Journal
of Sport Science 23 (10), 2098–2108. https://doi.org/10.1080/17461391.2023.2171906.
Farach-Carson, M.
C./Carson, D. D. (2007). Perlecan--a multifunctional extracellular
proteoglycan scaffold. Glycobiology 17 (9), 897–905. https://doi.org/10.1093/glycob/cwm043.
Feldmann, D. (2022).
Whole genome sequencing approach to identifying genetic risk factors
underlying anterior cruciate ligament injuries in a twin family study.
Faculty of Health Sciences, Department of Human Biology, 2022. Available
online at http://hdl.handle.net/11427/36615.
Feldmann, D. C./Rahim, M./Suijkerbuijk, M. A.
M./Laguette, M.-J. N./Cieszczyk, P./Ficek, K./Huminska-Lisowska, K./Häger,
C. K./Stattin, E./Nilsson, K. G./Alvarez-Rumero, J./Eynon, N./Feller,
J./Tirosh, O./Posthumus, M./Chimusa, E. R./Collins, M./September, A. V.
(2022). Investigation of multiple populations highlight VEGFA polymorphisms
to modulate anterior cruciate ligament injury. Journal of Orthopaedic
Research: Official Publication of the Orthopaedic Research Society 40 (7),
1604–1612. https://doi.org/10.1002/jor.25192.
Gibbon, A./Saunders, C. J./Collins, M./Gamieldien,
J./September, A. V. (2018). Defining the molecular signatures of Achilles
tendinopathy and anterior cruciate ligament ruptures: A whole-exome
sequencing approach. PloS One 13 (10), e0205860. https://doi.org/10.1371/journal.pone.0205860.
González, J.
R./Armengol, L./Solé, X./Guinó, E./Mercader, J. M./Estivill, X./Moreno, V.
(2007). SNPassoc: an R package to perform whole genome association studies.
Bioinformatics 23 (5), 644–645. https://doi.org/10.1093/bioinformatics/btm025.
Griffin, L.
Y./Albohm, M. J./Arendt, E. A./Bahr, R./Beynnon, B. D./Demaio, M./Dick, R.
W./Engebretsen, L./Garrett, W. E./Hannafin, J. A./Hewett, T. E./Huston, L.
J./Ireland, M. L./Johnson, R. J./Lephart, S./Mandelbaum, B. R./Mann, B.
J./Marks, P. H./Marshall, S. W./Myklebust, G./Noyes, F. R./Powers,
C./Shields, C./Shultz, S. J./Silvers, H./Slauterbeck, J./Taylor, D.
C./Teitz, C. C./Wojtys, E. M./Yu, B. (2006). Understanding and preventing
noncontact anterior cruciate ligament injuries: a review of the Hunt Valley
II meeting, January 2005. The American Journal of Sports Medicine 34 (9),
1512–1532. https://doi.org/10.1177/0363546506286866.
Gubbiotti, M.
A./Neill, T./Iozzo, R. V. (2017). A current view of perlecan in physiology
and pathology: A mosaic of functions. Matrix Biology: Journal of the
International Society for Matrix Biology 57-58, 285–298. https://doi.org/10.1016/j.matbio.2016.09.003.
Hayes, A.
J./Farrugia, B. L./Biose, I. J./Bix, G. J./Melrose, J. (2022). Perlecan, A
Multi-Functional, Cell-Instructive, Matrix-Stabilizing Proteoglycan With
Roles in Tissue Development Has Relevance to Connective Tissue Repair and
Regeneration. Frontiers in Cell and Developmental Biology 10, 856261. https://doi.org/10.3389/fcell.2022.856261.
Hermanussen,
M./Aßmann, C./Groth, D. (2021). Chain Reversion for Detecting Associations
in Interacting Variables-St. Nicolas House Analysis. International Journal
of Environmental Research and Public Health 18 (4). https://doi.org/10.3390/ijerph18041741.
Hewett, T. E./Myer,
G. D./Ford, K. R./Paterno, M. V./Quatman, C. E. (2016). Mechanisms,
prediction, and prevention of ACL injuries: Cut risk with three sharpened
and validated tools. Journal of Orthopaedic Research: Official Publication
of the Orthopaedic Research Society 34 (11), 1843–1855. https://doi.org/10.1002/jor.23414.
Huang, J./Li, X./Shi, X./Zhu, M./Wang, J./Huang,
S./Huang, X./Wang, H./Li, L./Deng, H./Zhou, Y./Mao, J./Long, Z./Ma, Z./Ye,
W./Pan, J./Xi, X./Jin, J. (2019). Platelet integrin αIIbβ3: signal
transduction, regulation, and its therapeutic targeting. Journal of
Hematology & Oncology 12 (1), 26. https://doi.org/10.1186/s13045-019-0709-6.
Hurd, W. J./Axe, M.
J./Snyder-Mackler, L. (2008). Influence of age, gender, and injury mechanism
on the development of dynamic knee stability after acute ACL rupture. The
Journal of Orthopaedic and Sports physical Therapy 38 (2), 36–41. https://doi.org/10.2519/jospt.2008.2609.
Iozzo, R. V. (2005).
Basement membrane proteoglycans: from cellar to ceiling. Nature Reviews.
Molecular Cell Biology 6 (8), 646–656. https://doi.org/10.1038/nrm1702.
Joseph, A. M./Collins, C. L./Henke, N. M./Yard, E.
E./Fields, S. K./Comstock, R. D. (2013). A multisport epidemiologic
comparison of anterior cruciate ligament injuries in high school athletics.
Journal of Athletic Training 48 (6), 810–817. https://doi.org/10.4085/1062-6050-48.6.03.
Kızılgöz,
V./Sivrioğlu, A. K./Aydın, H./Ulusoy, G. R./Çetin, T./Tuncer, K. (2019). The
Combined Effect of Body Mass Index and Tibial Slope Angles on Anterior
Cruciate Ligament Injury Risk in Male Knees: A Case-Control Study. Clinical
Medicine Insights. Arthritis and Musculoskeletal Disorders 12,
1179544119867922. https://doi.org/10.1177/1179544119867922.
Kobayashi,
H./Kanamura, T./Koshida, S./Miyashita, K./Okado, T./Shimizu, T./Yokoe, K.
(2010). Mechanisms of the anterior cruciate ligament injury in sports
activities: a twenty-year clinical research of 1,700 athletes. Journal of
Sports science & Medicine 9 (4), 669–675.
Lahiri, D. K./Nurnberger, J. I. (1991). A rapid
non-enzymatic method for the preparation of HMW DNA from blood for RFLP
studies. Nucleic Acids Research 19 (19), 5444. https://doi.org/10.1093/nar/19.19.5444.
Logerstedt, D.
S./Scalzitti, D. A./Bennell, K. L./Hinman, R. S./Silvers-Granelli, H./Ebert,
J./Hambly, K./Carey, J. L./Snyder-Mackler, L./Axe, M. J./McDonough, C. M.
(2018). Knee Pain and Mobility Impairments: Meniscal and Articular Cartilage
Lesions Revision 2018. The Journal of Orthopaedic and Sports physical
Therapy 48 (2), A1-A50. https://doi.org/10.2519/jospt.2018.0301.
Lohmander, L. S./Englund, P. M./Dahl, L. L./Roos, E.
M. (2007). The long-term consequence of anterior cruciate ligament and
meniscus injuries: osteoarthritis. The American Journal of Sports Medicine
35 (10), 1756–1769. https://doi.org/10.1177/0363546507307396.
Lulińska-Kuklik,
E./Leźnicka, K./Humińska-Lisowska, K./Moska, W./Michałowska-Sawczyn,
M./Ossowski, Z./Maculewicz, E./Cięszczyk, P./Kaczmarczyk, M./Ratkowski,
W./Ficek, K./Zmijewski, P./Leońska-Duniec, A. (2019). The VEGFA gene and
anterior cruciate ligament rupture risk in the Caucasian population. Biology
of Sport 36 (1), 3–8. https://doi.org/10.5114/biolsport.2018.78902.
Magnusson,
K./Turkiewicz, A./Hughes, V./Frobell, R./Englund, M. (2020). High genetic
contribution to anterior cruciate ligament rupture: Heritability ~69.
British Journal of Sports Medicine. https://doi.org/10.1136/bjsports-2020-102392.
Martinez, J. R./Dhawan, A./Farach-Carson, M. C.
(2018). Modular Proteoglycan Perlecan/HSPG2: Mutations, Phenotypes, and
Functions. Genes 9 (11). https://doi.org/10.3390/genes9110556.
Meeuwisse, W. (1994). Assessing Causation in Sport
Injury: A Multifactorial Model. Clinical Journal of Sport Medicine 4 (3),
166–170.
Mokone, G.
G./Schwellnus, M. P./Noakes, T. D./Collins, M. (2006). The COL5A1 gene and
Achilles tendon pathology. Scandinavian Journal of Medicine & Science in
Sports 16 (1), 19–26. https://doi.org/10.1111/j.1600-0838.2005.00439.x.
Motififard, M./Akbari
Aghdam, H./Ravanbod, H./Jafarpishe, M. S./Shahsavan, M./Daemi, A./Mehrvar,
A./Rezvani, A./Jamalirad, H./Jajroudi, M./Shahsavan, M. (2024). Demographic
and Injury Characteristics as Potential Risk Factors for Anterior Cruciate
Ligament Injuries: A Multicentric Cross-Sectional Study. Journal of Clinical
Medicine 13 (17). https://doi.org/10.3390/jcm13175063.
Murphy, D. F./Connolly, D. A. J./Beynnon, B. D.
(2003). Risk factors for lower extremity injury: a review of the literature.
British Journal of Sports Medicine 37 (1), 13–29. https://doi.org/10.1136/bjsm.37.1.13.
Murray, M. M./Martin,
S. D./Martin, T. L./Spector, M. (2000). Histological changes in the human
anterior cruciate ligament after rupture. The Journal of Bone and Joint
Surgery. American Volume 82 (10), 1387–1397. https://doi.org/10.2106/00004623-200010000-00004.
Posthumus,
M./September, A. V./O'Cuinneagain, D./van der Merwe, W./Schwellnus, M.
P./Collins, M. (2010). The association between the COL12A1 gene and anterior
cruciate ligament ruptures. British Journal of Sports Medicine 44 (16),
1160–1165. https://doi.org/10.1136/bjsm.2009.060756.
Rahim, M./Mannion,
S./Klug, B./Hobbs, H./van der Merwe, W./Posthumus, M./Collins, M./September,
A. V. (2017). Modulators of the extracellular matrix and risk of anterior
cruciate ligament ruptures. Journal of Science and Medicine in Sport 20 (2),
152–158. https://doi.org/10.1016/j.jsams.2016.07.003.
Ribbans, W.
J./September, A. V./Collins, M. (2022). Tendon and Ligament Genetics: How Do
They Contribute to Disease and Injury? A Narrative Review. Life 12 (5). https://doi.org/10.3390/life12050663.
Schaid, D. J./Sinnwell, J. P./Jenkins, G.
D./McDonnell, S. K./Ingle, J. N./Kubo, M./Goss, P. E./Costantino, J.
P./Wickerham, D. L./Weinshilboum, R. M. (2012). Using the gene ontology to
scan multilevel gene sets for associations in genome wide association
studies. Genetic Epidemiology 36 (1), 3–16. https://doi.org/10.1002/gepi.20632.
Seneviratne, A./Attia, E./Williams, R. J./Rodeo, S.
A./Hannafin, J. A. (2004). The effect of estrogen on ovine anterior cruciate
ligament fibroblasts: cell proliferation and collagen synthesis. The
American Journal of Sports Medicine 32 (7), 1613–1618. https://doi.org/10.1177/0363546503262179.
Shapiro, S. S./Wilk,
M. B. (1965). An Analysis of Variance Test for Normality (Complete Samples).
Biometrika 52 (3/4), 591. https://doi.org/10.2307/2333709.
Snaebjörnsson, T./Svantesson, E./Sundemo, D./Westin,
O./Sansone, M./Engebretsen, L./Hamrin-Senorski, E. (2019). Young age and
high BMI are predictors of early revision surgery after primary anterior
cruciate ligament reconstruction: a cohort study from the Swedish and
Norwegian knee ligament registries based on 30,747 patients. Knee Surgery,
Sports Traumatology, Arthroscopy: Official Journal of the ESSKA 27 (11),
3583–3591. https://doi.org/10.1007/s00167-019-05487-2.
Suijkerbuijk, M. A.
M./Ponzetti, M./Rahim, M./Posthumus, M./Häger, C. K./Stattin, E./Nilsson, K.
G./Teti, A./Meuffels, D. E./van der Eerden, B. J. C./Collins, M./September,
A. V. (2019). Functional polymorphisms within the inflammatory pathway
regulate expression of extracellular matrix components in a genetic risk
dependent model for anterior cruciate ligament injuries. Journal of Science
and Medicine in Sport 22 (11), 1219–1225. https://doi.org/10.1016/j.jsams.2019.07.012.
Suter, L. G./Smith,
S. R./Katz, J. N./Englund, M./Hunter, D. J./Frobell, R./Losina, E. (2017).
Projecting Lifetime Risk of Symptomatic Knee Osteoarthritis and Total Knee
Replacement in Individuals Sustaining a Complete Anterior Cruciate Ligament
Tear in Early Adulthood. Arthritis care & Research 69 (2), 201–208. https://doi.org/10.1002/acr.22940.
Widmyer, M. R./Utturkar, G. M./Leddy, H. A./Coleman,
J. L./Spritzer, C. E./Moorman, C. T./DeFrate, L. E./Guilak, F. (2013). High
body mass index is associated with increased diurnal strains in the
articular cartilage of the knee. Arthritis and Rheumatism 65 (10),
2615–2622. https://doi.org/10.1002/art.38062.
Willard, K./Mannion, S./Saunders, C. J./Collins,
M./September, A. V. (2018). The interaction of polymorphisms in
extracellular matrix genes and underlying miRNA motifs that modulate
susceptibility to anterior cruciate ligament rupture. Journal of Science and
Medicine in Sport 21 (1), 22–28. https://doi.org/10.1016/j.jsams.2017.08.017.